Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Зона неморальных, или летнезелёных лесов - Широтная зональность - Биогеография суши
Биомасса (сухая, надземных и подземных частей) растений в климаксных дубравах Quercus robur превышает 400 т/га, на долю травяного покрова приходится 0,06% (Вальтер, 1975а). А.Е. Родин и Н.И. Базилевич (1965) указывают для молодого, 200летнего ельника-зеленомошника в Архангельской области 341 т/га. Листья деревьев образуют 1—2% биомассы дерева, хвоя 5—7% (Родин, Базилевич, 1965). Позвоночные образуют биомассу 0,01—0,03 т/га, беспозвоночные (в основном дождевые черви) — до 1 т/га и более. Добавление в почву нафталина, не влияющего существенно на грибы и бактерии, но губящего беспозвоночных, снижало скорость переработки подстилки в шесть раз (Вальтер, 1975а). Чистая продуктивность (т/га сухого вещества в год) букового леса в Дании та же (13,5), что и тропического дождевого леса на Береге Слоновой Кости (13,4), тогда как брутто-продуктивность (включая расходы растений на дыхание) вдвое меньше (23,5 и 52,5, соответственно), что объясняют более высокой температурой в тропиках. Родин, Базилевич (1965) указывают годовой прирост в 220-летней дубраве в 6,5 т/га, реальная продуктивность существенно выше, поскольку эти данные не учитывают потребление продукции фитофагами в течение года. Содержание гумуса в почвах составляет 1—70 т/га (Вальтер, 1968). В дубовых лесах лесостепи млекопитающие потребляют 0,3 т/га фитомассы. Продолжительность жизни деревьев климаксных ассоциаций достигает многих сотен лет. Простые подсчёты показывают, что при возрасте деревьев в лесу в 2000 лет для нормального возобновления достаточно, чтобы на 1 га вырастало 1 дерево в год. И, хотя обычно возраст деревьев гораздо меньше, обилие подроста может быть меньше фиксируемого стандартными геоботаническими методами и лес будет интерпретирован как отмирающий.
Существенную роль в почвах умеренного пояса, особенно в лесных ассоциациях играют дождевые черви Lumbricidae, роль дождевых червей в других климатических зонах относительно меньше. Также для умеренного пояса характерны кроты Talpidae.
Деление деревьев на “широколиственные” и “мелколиственные” довольно условно. Обычно к широколиственным породам относят из растущих в средней полосе России дуб, граб, бук, липу, клён платанолистный, вяз, а к мелколиственным — берёзу и осину. Куда отнести ольху и черёмуху по размерам их листьев неясно, поскольку их листовые пластинки имеют промежуточный размер, ольху обычно относят к мелколиственным, а черёмуху — к широколиственным. В действительности, деревья, которые считают широколиственными имеют не столь уж большие листья. Так, если сравнить листья западноевропейской липы крупнолистной Tilia platyphillos с растущей в России T. cordata — липой мелколистной, понимаешь, почему они так названы.
Примеры климадиаграмм районов произрастания широколиственных лесов показаны на Рис. 3.23. Эдификаторы лесных климаксных ассоциаций: листопадные дубы: в Евразии обыкновенный Quercus robur, пушистый Q. pubescens, скальный Q. petraea, монгольский Q. mongolica, расширенный Q. dilatata, зубчатый Q. dentata, в Северной Америке Гаррьяна Q. garryana, Гамбела Q. gambeli, красный, или северный Q. rubra, каштановый Q. prinus. В неморальных лесах высоко разнообразие эфемероидов — растений вегетирующих до распускания листвы деревьев, когда они дают до 100% проективного покрытия при плотности поселений до 10 000 экз./кв. м. На их смену летом приходит разнотравье, образованное высокими (до 1,5 м) травами: злаками, сложноцветными, лилейными и др. Напочвенный покров отсутствует, поскольку скорость накопления подстилки больше, чем скорость роста мхов и лишайников. Хорошо развиты ярусы кустарников и невысоких деревьев.
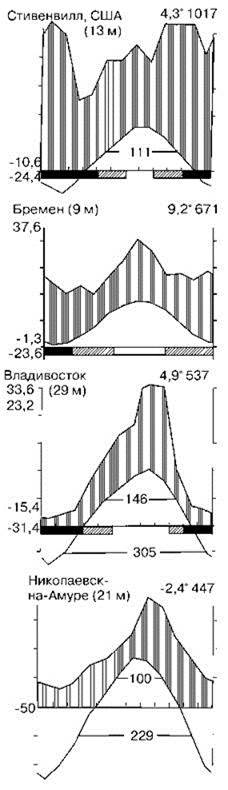
Рис. 3.23. Примеры климадиаграмм районов широколиственных лесов в Северном полушарии. По Вальтер (1975а,б) c дополнениями. См. также Рис. 3.8.
Изучение биологии сохранившихся видов млекопитающих и птиц северной Евразии показывает, что видов, приуроченных к крупным массивам лесов, климаксных и предклимаксных, очень мало, большинство предпочитает опушки, мозаичные биотопы и т. п. Ю.И. Чернов (1975), проанализировав данные по широко распространённым в северной Евразии видам млекопитающих, птиц и беспозвоночных, пришёл к выводу, что наибольшую плотность поселений и наибольшую эвритопность (которую он определял как площадь, занятую лакунами кружева ареала) эти виды имеют в лесостепи. Изучение биологии растений лесного пояса показало, что в сплошном лесу “только незначительная часть этих видов способна к... устойчивому самоподдержанию популяций” (Смирнова, 2004: 25). Для нормального возобновления растений необходима достаточно высокая освещённость, которой нет в сомкнутом древостое. Растения “существуют или благодаря антропогенному поддержанию.., или вследствие сочетания антропогенных и природных механизмов поддержания популяций” (Смирнова, 2004: 25). Это дополнительно подтверждает отсутствие обширных сплошных лесных массивов в совсем недавнем прошлом. Выпадение мегафауны из экосистем и возникновение крупных массивов лесов должно снизить и численность видов открытых пространств и опушек. Вероятно, это и привело к вымиранию части из них, в том числе и мелких птиц и млекопитающих, на которых человек не охотился, как случилось в Северной Америке. Поэтому нельзя рассматривать вымирание мелких птиц в послеледниковый период, как свидетельство несвязанного с человеком вымирания мегафауны, как это делает J.P. Grayson(1977). Полного вымирания мелких позвоночных и беспозвоночных, связанных с экотонами и ранними (безлесными) сукцессионными стадиями растительности, всё же обычно не происходит: для поддержания жизнеспособных популяций мелких животных, как правило, хватает и резко сократившейся площади полян, опушек и т. д.
Одним из основных видов-эдификаторов зон летнезелёных лесов и тайги был мамонт. В поддержании мозаичности велика была роль других видов мегафауны. Большинство из них было специфично для Евразии или Северной Америки. Некоторое представление о том, как выглядела Голарктика до прихода человека, дают африканские саванны. Следует учитывать следующие моменты, позволяющие допустить, что роль крупных травоядных млекопитающих и, возможно, их продуктивность в умеренном поясе были даже выше, чем в саванне:
— различия в продолжительности вегетационного периода незначительны: на большей части территории саванн и Голарктики — порядка полугода. В Голарктике его ограничивает температура, в саваннах — увлажнение.
— различия в структуре фитофагов. В африканских саваннах более половины первичной продукции изымают термиты, велика роль растительноядных насекомых, в частности прямокрылых. В мамонтовых СС не было термитов (как нет их и сейчас в умеренном поясе), роль саранчовых существенно ниже. Поэтому мегафауне Голарктики доставалась существенно большая часть первичной продукции, чем ныне в тропиках.
Освободившиеся в результате вымирания крупных фитофагов пищевые ресурсы стали потреблять грызуны, что привело к существенному увеличению их роли в экосистемах (Абатуров, 1984; Kalandadze, Rautian, 1995).
Значения биомассы и продуктивности различных растительных ассоциаций разных зон даны в Табл. 3.4.
Естественное состояние СС суши — мозаика лесных и безлесных участков. Однако безлесные ассоциации столь широко распространены, что их выделяют в особые типы зональной растительности: тундры, степи, пустыни и саванны. Об антропогенной природе саванн сказано выше. Рассмотрим теперь природу безлесных ландшафтов зоны летнезелёных лесов — степей.
Степи. Безлесие степей имеет две основные причины. Устранение древесных ассоциаций из первично мозаичного ландшафта следует считать антропогенным, связанным с вырубанием деревьев и их уничтожением пожарами. С другой стороны, хотя для сплошного леса в безлесных участках степи влаги не хватает, отдельные деревья вполне могли бы там существовать. Однако во флоре умеренной зоны отсутствуют виды деревьев, способные расти одиночно на фоне сомкнутого травостоя, образуя саванноподобные ландшафты, а есть лишь виды, образующие сомкнутые леса.
Табл. 3.4. Биомасса и продуктивность (сухого вещества т/га) разных сообществ Евразии. По Абдурахманову и др. (2003).
|
ландшафт |
фито масса, т/га |
продукция |
годовой Р/В |
зоомасса, кг/га |
|
тропические влажные вечнозелёные леса |
350-700 |
6-50 |
0,01-0,1 |
1000 |
|
тропические листопадные леса |
200-250 |
10 |
||
|
тропические листопадные редколесья |
<150 |
|||
|
тропические листопадные кустарники |
<100 |
|||
|
саванны |
50-150 |
5-15 |
0,1 |
100-130 |
|
субтропические жестколистные леса и кустарники |
50 |
10 |
0,2 |
|
|
степи и прерии |
10-150 |
5-30 |
0,2-0,5 |
10-50 |
|
широколиственные леса умеренного пояса |
400-500 |
10-50 |
до 1000 |
|
|
бореальные хвойные леса |
350-400 |
8-10 |
||
|
тундры |
1-50 |
1-5 |
0,1-0,2 |
Именно поэтому граница между лесными и степными участками мозаичных ландшафтов умеренных широт столь резка, в отличие от плавного перехода между ними в тропиках и субтропиках. Лишь в Северной Америке, где безлесные климаксы сформировались раньше, чем в Евразии, некоторые виды деревьев способны образовывать некоторое подобие саванны, например дуб крупноплодный Quercus macrocarpa. Корень взрослого дерева проникает на глубину свыше 5 м, длина боковых корней достигает 18 м, а толстая кора позволяет дубу переживать пожары.
Безлесные ассоциации существенно изменились даже за последние несколько столетий. Развитие земледелия и скотоводства привело к сильной эрозии почв, развитию дренирующей сети оврагов и балок и врезу русел рек — всё это способствовало обезвоживанию степи и колоссальной её эрозии. Такие же процессы происходили повсеместно в других районах. О масштабах эрозии в прериях Северной Америки наглядно свидетельствует случай, описанный Джаредом Даймондом (2008: 678). В штате Айова (одного из основных сельскохозяйственных районов США) в XIX в. была построена церковь. В момент постройки её двор располагался на уровне остальных домов. Всего за 150 лет окружающая её местность стала ниже на 3 метра и ныне церковь возвышается над окружающими полями как маленький остров. Сходных масштабов эрозия достигала на целинных землях СССР после их распашки в 1954—1961 гг. В Южной Америке и в середине ХХ в. в юго-западной Африке “вместо прежних обширных зарослей высоких злаков вдоль рек сегодня тянутся безжизненные песчаные пространства” (Вальтер, 1975: 97).
Пустыни. Большая часть пустынь Земли имеет антропогенный характер, естественными являются лишь немногие пустыни: береговые пустыни районов апвеллингов, пустыни горных долин (типа Гоби). Об антропогенном характере Сахары сказано выше. О пустынях Средней и Центральной Азии см. Вальтер (1975б, стр. 200 и далее). Нет естественных пустынь и в Австралии (см. ниже).
“Опустынивание происходит в результате действия косвенных факторов, обусловливающих неустойчивое использование скудных природных ресурсов местными землепользователями” (Оценка экосистем..., 2005: 10).